Гибридные РНК могут инициировать перестройки хромосом в клетках человека
При многих типах рака клетки приобретают характерные перестройки хромосом. Часто это приводит к слиянию частей двух разных генов. В результате транскрипции и трансляции на таких объединенных генах образуются гибридные РНК и белки, работа которых как раз и вызывает аномалии в поведении раковых клеток. Однако время от времени врачам попадаются случаи, когда гибридные РНК присутствуют у пациентов, хотя собственно хромосомной перестройки нет. Это объясняется явлением транс-сплайсинга – пересшивания частей двух нормальных молекул РНК. Американские ученые в серии прямых экспериментов продемонстрировали, что транс-сплайсинг может предварять и инициировать возникновение соответствующей перестройки хромосом. В основе механизма лежит прямое взаимодействие между гибридной РНК и геномной ДНК.
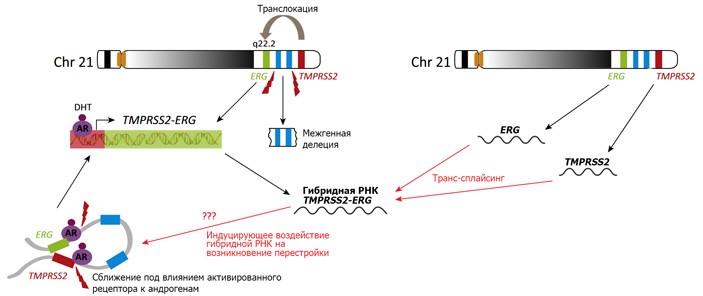
Перестройки хромосом характерны для многих видов рака. Перестройка возникает тогда, когда в двух разных хромосомах (или в разных участках одной и той же хромосомы) одновременно появляются двуцепочечные разрывы. При репарации (починке) таких разрывов может произойти ошибочное сшивание неподходящих друг к другу фрагментов. Если разрывы произошли внутри кодирующих белки генов, то пересшивание фрагментов ДНК приводит к появлению гибридных генов, в которых соединены куски нормальных. Гибридные продукты таких перестроек часто оказываются онкогенами: меняют поведение клеток с нормального на злокачественное, в результате чего и развивается рак.
Менее известно, что такие же гибридные продукты иногда обнаруживаются и у пациентов, в клетках которых отсутствует хромосомная перестройка. В этих случаях гибридизация двух генов происходит не на уровне ДНК, а на уровне РНК, при сплайсинге. Обычно сплайсинг происходит в пределах единой цепочки РНК, но здесь речь идет о так называемом транс-сплайсинге – сшивании частей двух разных молекул РНК.
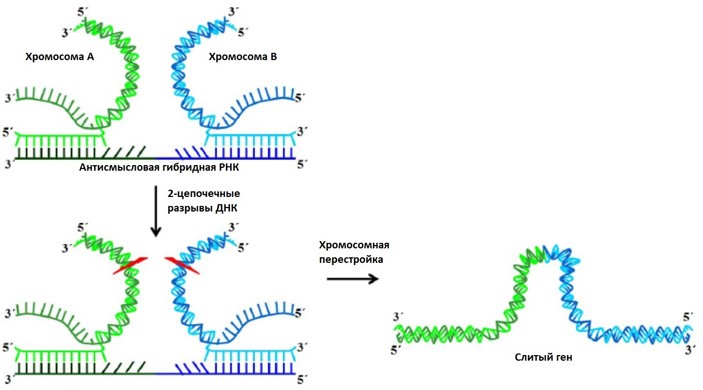
Оба механизма создают порой одинаковые или очень похожие гибридные продукты, обнаруживаемые в клетках пациентов (например, гибридный белок AML1-ETO при остром миелоидном лейкозе, состоящий из фрагментов нормальных генов AML1 и ETO, локализованных, соответственно, на 21 и 8 хромосомах). В связи с этим некоторыми авторами ранее была высказана гипотеза, предполагающая, что первоначально в клетке могут появляться РНК-продукты транс-сплайсинга, которые затем каким-то образом провоцируют появление соответствующей хромосомной перестройки. Эту гипотезу коротко называют «телега впереди лошади», так как исходное изменение в молекуле РНК переходит в изменение ДНК, а не наоборот (как это обычно предполагается в рамках «догмы» молекулярной биологии, см. схему на рисунка 1). Однако прямой демонстрации реальности такого сценария до сих пор не было.
Провести необходимые эксперименты для окончательного подтверждения или опровержения изложенной гипотезы взялся коллектив авторов из Медицинского колледжа Бейлора (Хьюстон, Техас, США). В качестве исследовательских моделей они ориентировались на две хромосомные перестройки, особенно часто встречающиеся при раке простаты у человека. Первая, внутрихромосомная (затрагивает хромосому 21), связана с появлением гибридного гена TMPRSS2-ERG. Вторая, межхромосомная (объединяющая фрагменты 21 и 7 хромосом), образует гибридный ген TMPRSS2-ETV1. Эксперименты проводились на линии клеток простаты человека LNCaP. Сама по себе эта линия не содержит ни первой, ни второй из описанных транслокаций.
Клетки линии LNCaP чувствительны к андрогенам, что в целом характерно для ткани предстательной железы. В более ранних работах было обнаружено, что вероятность хромосомных перестроек в клетках рака простаты повышается в ответ на присутствие андрогенов. Гормон проникает в клетки и активирует стероидные рецепторы. Активированные рецепторы, в свою очередь, проникают в ядро и активируют транскрипцию генов, зависимых от андрогенов. Предполагается, что эти события вызывают перераспределение материала хромосом в ядре и сближение участков хромосом, на которых располагаются такие гены. Дело в том, что в ядре наших клеток существуют специфические компартменты – транскрипционные фабрики, где сконцентрированы молекулы фермента РНК-полимеразы, и именно в этих компартментах и осуществляется транскрипция. Транскрипционные фабрики не отделены от окружающего пространства ядра особой мембраной, но, тем не менее, имеют вполне конкретную локализацию и очерченные границы. Такой механизм повышает вероятность возникновения хромосомных перестроек именно между активными генами: ведь если в двух таких участках возникнут разрывы, то оборванные концы оказываются рядом, и есть некоторая вероятность их случайного объединения в ходе репарации по пути негомологичного соединения концов. С учетом всего вышесказанного, эксперименты на клетках линии LNCaP проводили с добавлением в культуральную среду дигидротестостерона (DHT).
Идея заключалась в том, чтобы методами генетической модификации ввести в клетки минигены, кодирующие гибридные РНК двух зависимых от тестостерона генов, и проверить, не последует ли за этим появление заранее спланированных хромосомных перестроек.
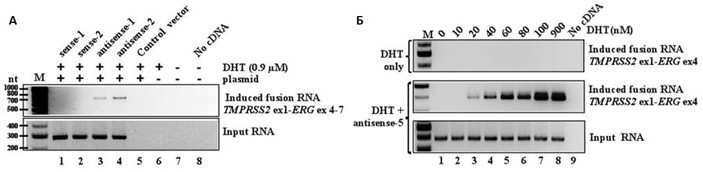
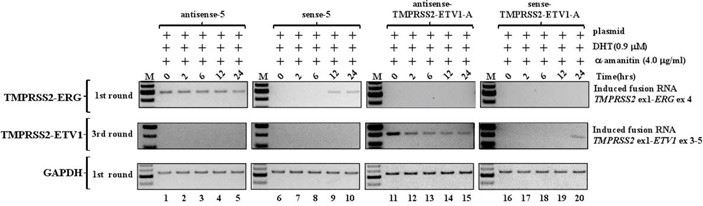
В первой серии экспериментов предприняли попытки инициировать транслокацию TMPRSS2-ERG. Такая мутация встречается примерно в половине всех случаев рака простаты. Исследователи создали несколько вариантов векторных конструкций, в каждой из которых был встроен участок для обеспечения экспрессии небольшого фрагмента (около 200 пар нуклеотидов) гибридной РНК, соответствующей смысловой или антисмысловой цепи гибридного гена. Часть каждой РНК соответствовала первому экзону гена TMPRSS2, другая часть – четвертому экзону гена ERG: именно между этими экзонами чаще всего располагаются точки разрыва-слияния при раке простаты. После проведения генетической модификации клетки культивировали, и спустя несколько дней выделяли из них РНК и анализировали ее методом ПЦР с использованием двух разных пар праймеров. Первая пара выявляла присутствие укороченных гибридных РНК, которые должны были нарабатываться в клетках после искусственной модификации. Другая пара праймеров могла сработать только на полном гибридном гене, которого, как уже сказано, в линии клеток LNCaP изначально нет. В случае появления ожидаемой хромосомной перестройки и транскрипции гибридного гена, в ПЦР со второй парой праймеров должен был получиться фрагмент длиной около 700 нуклеотидов.
Некоторые конструкции действительно сработали: в модифицированных клетках появлялась полная гибридная РНК! Тут стоит обратить внимание, что транслокация возникала лишь в небольшой доле клеток, подвергшихся модификации (0,1% или менее), но результат стабильно воспроизводился при повторных экспериментах. Неожиданным оказалось то, что при прочих равных условиях сработали только те векторы, в которых искусственный ген кодировал антисмысловые (antisense) последовательности, в то время как смысловые (sense) гибридные РНК не индуцировали хромосомной перестройки (сравните дорожки 1, 2 и 3, 4 в геле на рисунке 2, А). Клетки, модифицированные генами для антисмысловых РНК, демонстрировали явное повышение эффективности индукции транслокации по мере повышения концентрации добавляемого DHT: мутации не возникало вовсе, если DHT содержался в концентрации менее 20 нМ (рисунке 2, Б, средний гель). Также мутации не возникали при любых концентрациях DHT, но в отсутствие генетической модификации (рисунке 2, Б, верхний гель).
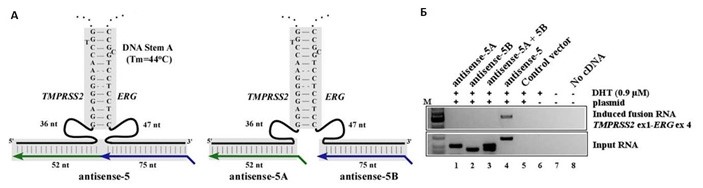
Множественные испытанные версии минигенов антисмысловых РНК оказались существенно различающимися по способности инициировать перестройку хромосом. Анализ последовательностей позволил выяснить, что результат можно предсказать по выявлению небольших участков микрогомологии вблизи области комплементарного присоединения антисмысловой РНК (длиной между 10 и 30 нуклеотидов). Чем более термодинамически стабильная «шпилька» формируется между нитями ДНК, тем с большей вероятностью инициируется хромосомной перестройки. Также критичным для стимуляции мутации оказалось одновременное присутствие в антисмысловой РНК областей комплементарности с обоими участками (в области гена TMPRSS2 и гена ERG). Транслокации не происходило, если миниген гибридной антисмысловой РНК подменяли двумя «половинчатыми» генами, каждый из которых комплементарен участку только гена TMPRSS2 или только гена ERG рисунок 3.
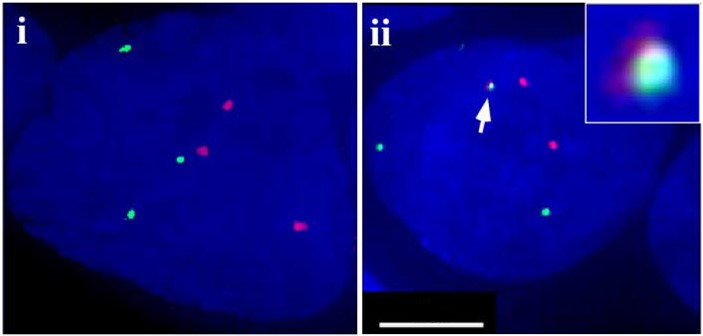
Для другого гибридного гена TMPRSS2-ETV1 результаты оказались очень похожими при том, что в этом случае перестройка затрагивает одновременно две разных хромосомы: 21 и 7. То, что хромосомная перестройка действительно имела место в некотором количестве экспериментальных клеток, было дополнительно подтверждено методом флуоресцентной гибридизации in situ (FISH) – двухцветной окраски хромосом, которая позволила увидеть в части клеток слитую хромосому, окрашенную одновременно двумя цветами рисунок 4. Перестройка была обнаружена в 0,91% от примерно 3000 просмотренных клеток, экспрессирующих гибридную антисмысловую РНК. Среди популяции клеток без экспрессии гибридных РНК перестройку не содержала ни одна (из более 600 просмотренных клеток).
Исходя из полученных результатов можно предположить, что функция гибридной РНК в инициации хромосомной перестройки сводится к удержанию двух удаленных участков на одной и той же или на разных хромосомах в тесном контакте. Тем самым создаются максимально благоприятные условия для случайного ошибочного сшивания концов в случае возникновения разрыва в нити ДНК (схема представлена на рисунке 5). Поскольку появление разрыва, как и ошибочное сшивание концов – дело случая, вполне ожидаемо, что перестройка будет возникать лишь в некоторой (небольшой) доле клеток, экспрессирующих гибридные РНК.
После всего сказанного остается неясным еще один вопрос: почему индукторами перестроек оказались только антисмысловые РНК, но не смысловые? Ведь теоретически сближение участков нити ДНК могло осуществляться как теми, так и другими? Было предположино, что смысловые РНК просто не могут удержаться на нити ДНК вследствие того, что их «сталкивают» проходящие по ДНК транскрипционные машины. Для проверки этого предположения был применен ингибитор транскрипции (альфа-актинин), который значительно замедляет продвижение транскрипционных комплексов. И действительно, в таких условиях смысловые РНК также стали работать как индукторы транслокации. Причем индуцирующее действие молекул РНК и в случае антисмысловых, и смысловых РНК было специфичным: последовательности инициировали транслокации только того типа, которому эти РНК соответствовали по последовательностям. Сам по себе альфа-актинин никакого инициирующего действия на возникновение перестроек не оказывал рисунок 6.
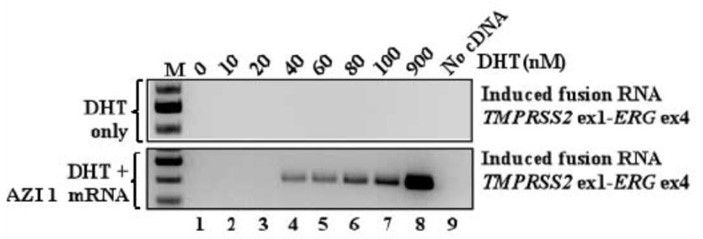
Но до сих пор речь шла исключительно об искусственных конструкциях. Между тем изначальной идеей было найти внутренний РНК-индуктор, который обуславливает столь частое возникновение однотипной транслокации (в частности, TMPRSS2-ERG) в клетках рака простаты. Поскольку было уже примерно понятно, что нужно искать, исследователи просканировали библиотеки некодирующих РНК (нкРНК), экспрессирующихся в ткани предстательной железы и действительно обнаружили то, что искали: в составе длинной некодирующей РНК, AZI1, выявилась потенциальная область комплементарного связывания с подходящими участками генов TMPRSS2 и ERG. Искусственное введение в клетки гена, обеспечивающего экспрессию только этого небольшого участка, как и предполагалось, приводило к появлению клеток с транслокацией рисунок 7. Тот же результат получали и при искусственной стимуляции экспрессии внутриклеточного гена AZI1. Между тем, в ходе экспериментов обнаружилось, что экспрессия гена AZI1 положительно зависит от концентрации андрогена (в данном случае, DHT).
Таким образом, впервые было установлено, что молекулы РНК действительно при определенных дополнительных условиях могут предопределять области наиболее вероятного появления хромосомных перестроек в клетках млекопитающих.
Источник:
Sachin Kumar Gupta, Liming Luo, Laising Yen. RNA-mediated Genomic Arrangements in Mammalian Cells // 2017. DOI: 10.1101/151241.
© Новиков Роман Игоревич, младший научный сотрудник лаборатории эндокринологии и биохимии.
Добавить комментарий